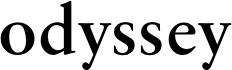



Introduction
Finding successful predictors and links between mating behavior and evolution can be taxing. Certain outliers like humans, some elephant groups, and hydras make it difficult to trace behavioral traits, like monogamy, back to a common ancestor. Current mating behavior descriptors include Adult Sex Ratio (ASR), Operational Sex Ratio (OSR), male/female Reproductive Success (RS), amount of sexually dimorphic characteristics, the amount of male-male competition, amount of female-female competition, and various others. These descriptors are often correlated with traits and can be used as predictors for a species proclivity toward monogamy, amount of sexual dimorphism, and even male-male competition(1.)
ASR represents the number of sexually mature males to females. ASR had been known to correlate with female distribution patterns, mortality, and maturation. A species’ OSR is the amount of sexually active males to sexually participating females (8). Although very similar, the ASR describes males and females that do mate while the OSR describes males and females that might mate. OSR tends to miss males that have reached sexual maturity “early,” as it only looks at those individuals who have visibly participated in mating during a certain observed period. Using ASR to describe a population tends to prevent miscounting, typically females can be over or undercounted in an OSR measurement because of menopause, concealed ovulation, due to individuals that reach those stages too early or too late (8). Female spacing behavior is a major mating strategy driver. It has been observed to be key in explaining monogamous mammalian behavior(2,4). In voles, a well-known monogamous mammalian species, female spacing behavior is one of the only traits that provides validity to the evolutionary advantage of monogamy as this species choice of mating strategy.
Hypothesis
In most primates exaggerated sexual dimorphism and high instances of male-male competition correlate with a high OSR (8). Humans have a higher OSR than expected for a species with little sexual dimorphism and low male-male competition even in hunter-gatherer societies. If OSR fails to predictably correlate with known measurements of sexual dimorphism/frequency of male-male competition then other factors such as visible ovulation resource distribution, resource inheritance patterns, ASR, need to be considered. To make better predictions about population extinction probabilities (a common use of OSR) and add to evolutionary history of humans, primates generally, and monogamous mammals overall.
The Proposal
It seems primate researchers prefer to report the OSR of their specimen and ignore the ASR (1). Future analyses of data on mating strategy correlates for all species might include ASR analysis, OSR analysis, and detailed examination of known correlates. So far, analyses seems inconsistent or vary from species to species. This inconsistency is an accepted phenomenon often attributed to differences between species energy requirements.
However, there are visible connections and unifying factors alongside OSR and ASR which might make mating behavior research more consistent and progress the field so that research might continue to unravel why very different organisms have similar mating systems. Instead of using sex ratios to predict mating behaviors examining an organism’s resource distribution, or female dispersal patterns may be a better indicator. Since resource distribution and female spacing behavior are used to explain species that do not behave in predicted patterns. For example, in the human model OSR falls short because humans (8), unlike most other primates have concealed ovulation and the females typically live well beyond their fertile years. These factors obscure the analyses of their OSR and I am unsure if human ASR has even been calculated due to the bias in primate research to ignore ASR and to equate OSR and ASR.




 Santa and Mrs. Claus
StableDiffusion
Santa and Mrs. Claus
StableDiffusion
 R2D2 and C3PO
StableDiffusion
R2D2 and C3PO
StableDiffusion
 Harry Potter and Ronald Weasley
StableDiffusion
Harry Potter and Ronald Weasley
StableDiffusion
 Mickey and Minnie Mouse
StableDiffusion
Mickey and Minnie Mouse
StableDiffusion
 Captain Kirk and Spock
StableDiffusion
Captain Kirk and Spock
StableDiffusion
 Sherlock and Watson
StableDiffusion
Sherlock and Watson
StableDiffusion
 Mario and Luigi
StableDiffusion
Mario and Luigi
StableDiffusion
 mr and mrs potato head
StableDiffusion
mr and mrs potato head
StableDiffusion
 Barack and Michelle Obama
StableDiffusion
Barack and Michelle Obama
StableDiffusion
 Netflix and chill
StableDiffusion
Netflix and chill
StableDiffusion
 Chip n Dale
StableDiffusion
Chip n Dale
StableDiffusion
 George and Martha Washington
StableDiffusion
George and Martha Washington
StableDiffusion
 Jesus & Peter
Jesus & Peter Marge and Homer Simpson
StableDiffusion
Marge and Homer Simpson
StableDiffusion
 Bonnie & Clyde
i.pinimg.com
Bonnie & Clyde
i.pinimg.com










